![]() 以下の文書は「自分の論文を自分で解説した」ものである。
以下の文書は「自分の論文を自分で解説した」ものである。
理化学研究所のホームページ で、よく「プレスリリース」が発表されている。 研究成果が一般の人にもわかりやすく、しかも間違いなく紹介されておりすばらしい。そこで、それを真似て私もここに「プレスに載らないプレスリリース」を出してみる。「プレスリリース風」に書いてみると実際の論文とはずいぶんと雰囲気の違うものになった。しかし同じ植物生理学者の間でも分野が異なると研究内容が理解できなかったりするのだから、このような形式で成果を表現することが必要なのかもしれない。
最近「メタボリックシンドローム」が注目されているが、私のこの研究結果は「植物のメタボリックシンドローム」と言えなくもないので、それをタイトルに書いてみた。人間では基礎代謝が異常になることは病気の元である。だが、植物ではかえって有用な産物(アミノ酸など)の量が増えて、人間にとっては都合がよいこともある。また、植物は特別な器官において、大量の油脂や蜜を作る。植物は代謝システムをうまく組織・時期特異的に制御する仕組みを構築することで、植物自身にとって有用な産物を作り出している。
微生物でも基礎代謝の異常から同様な現象が見られるようで、普遍的なことらしい。
Alterations of glucose metabolism in Escherichia coli mutants defective in respiratory-chain enzymes. Kihira C, Hayashi Y, Azuma N, Noda S, Maeda S, Fukiya S, Wada M, Matsushita K, Yokota A. J Biotechnol. 2012 Apr 30;158(4):215-23. PMID: 21740932 北海道大学農学研究院 横田教授のグループの成果 http://www.agr.hokudai.ac.jp/biseibutsu/ja/info.html
また植物には、環境ストレスや遺伝的な変異で代謝機能が低下しても、代謝経路全体を可塑的に変動させることで補償してしまう能力がある(代謝の可塑性)。動物でもそういう能力はあるだろう。しかし植物はそれがもっと強くて、厳しい条件でもしぶとく生きていけるようになっているらしい。
―――― ここから ――――
― 有用代謝産物増産技術の応用へ ―
平成18年7月
◇ポイント◇
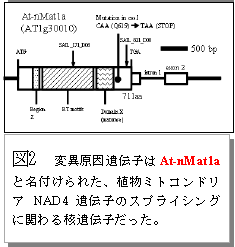
国立大学法人広島大学は、植物のミトコンドリアグループIIイントロンをスプライスする機能を持つ核ゲノム遺伝子 (At-nMat1a) に変異が生じると、セルロースを含む有用物質の蓄積量が変化することを発見しました。生物圏科学研究科に所属する中川らによる研究成果です。
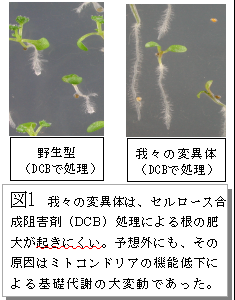
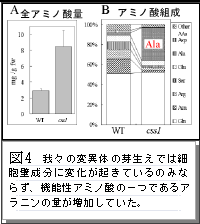
植物はセルロース、デンプン、脂質などの様々な有用代謝産物を生産します。これらの有用物質の生産には基礎的な炭素、窒素の代謝が本質的な役割を果たしています。植物細胞の基礎代謝は細胞質、葉緑体、ミトコンドリアの三者に局在する酵素群が巧妙に連携して進行しています。今回の研究は、この基礎代謝の連携を調節するメカニズムの一端を解明したものです。動物と異なり、植物のミトコンドリアゲノムにはイントロン(グループII)があります。研究グループは有用物質であるセルロースの合成機構を探る目的で、セルロース合成阻害剤処理による形態変化がおきにくい変異体 (図1) を単離し分析していました。その結果、その変異体ではミトコンドリアゲノムのイントロンをスプライスする因子に変異があり (図2, 3) 、ミトコンドリアの代謝機能に影響が出ていることを見いだしました。さらに、それがミトコンドリア外で行われる有用物質生産にも間接的に影響を与えている (図4) ことも明らかにしました。植物のミトコンドリア機能を RNA レベルで制御する遺伝子に、細胞全体の有用物質生産能の制御を行う機能があることを示した画期的な研究成果です。ミトコンドリアの機能制御を人為的に可能にすることで植物の有用代謝産物生産制御技術への応用が期待されます。
本研究成果は、日本植物生理学会が発行する英文国際誌「Plant Cell Physiology」の2006年6月号にすでに掲載されています。一部の人々からは、ある程度の評価を受けています。 証拠:http://www.researcherid.com/rid/C-9152-2009 このリスト もう一つの被引用リスト
(問い合わせ先) 中川 直樹 naka@hiroshima-u.ac.jp
(報道担当) なし


この成果については、すでに 「広島大学リエゾンフェア2005 in 東広島」という催しでも発表させていただいている。
その時に出展したポスターを以下に掲載する。
タイトル: 機能性アミノ酸、アラニンの蓄積量が増加したシロイヌナズナ(アブラナ科植物)の芽生え(スプラウト)の発見
このポスターを発表した後、一度カイワレダイコンを作っている会社から電話がかかってきた。しかしその後何の音沙汰もない。

私がシロイヌナズナで見つけた変異体に相当する、ダイコンの変異体をみつけるのは難しいだろう。そうしなくても、育成、処理条件を工夫することによって他の植物でも同様な「代謝改変効果」を得ることができるかもしれない。すでに低温条件を用いた技術が実用化されている。
例えば育成の最後期に低温ストレス等にさらすことによって糖などの代謝を変化させる。海産物では「氷温貯蔵」という低温処理を行うことで、生体に含まれるうまみ成分を増加させることが成功しているそうである。「雪の下でキャベツをおいしくする技術」が、既に実用化されているそうである。検索すると北海道で多く作られているようである。貝割れ大根でもうまくいくかもしれない。しかし低温にするのにコストがかかるのでは困る。工夫の余地があるのだろう。研究の段階では遺伝子組み換えや変異体の単離と分析など様々な手法を用いることができる。それによってわかった、できるようになったことを実用に生かす際は、研究で行ったことをコストが安く世の中に受け入れられやすい方法で代用する方法を改めて開発しないといけないのだろう。
最近になってやっと気が付いたのだが、この論文で出た結果は、「生物でも、”バイオエネルギー、バイオマス”などと言い出すと、エネルギー保存則、熱力学による制限から逃れることはできないことがはっきりしてくる」ということを示している。
Cellulose synthesis and cell expansion are regulated by different mechanisms in growing Arabidopsis hypocotyls. Ivakov AA, Flis A, Apelt F, Fuenfgeld M, Scherer U, Stitt M, Kragler F, Vissenberg K, Persson S, Suslov D.Plant Cell. 2017 May 26. pii: tpc.00782.2016. doi: 10.1105/tpc.16.00782. PMID: 28550150
という論文では、セルロースの合成はセルロース合成酵素の細胞内輸送で制御される、そしてそれは炭素代謝機能の影響を大きく受けると言うことを示しているらしい。まだよく読んでいない。
この論文に出てくる css1 変異体の種子がほしい方は、私あてにメールを送ってください。
このデータを得るのに大変な時間と労力を使ったが、さらにそれから論文として発表できるまでに、1年と5ヶ月以上かかっている(よく考えるとそれくらいかかるのが当たり前のような気もする)。
変異体と、その原因遺伝子の単離から仕事を進めるのは強力な手法だが、「運試し、運次第」という側面もある。ある重要な遺伝子を見つけたいとしても、その遺伝子に機能的に重複した別の遺伝子が存在していると変異体としては見つかりにくくなる。見つかるべき遺伝子が複数あっても、その中のあるものだけが高頻度で出てくる(バイアス)こともありうる。また遺伝子を単離しても機能がわからないとか、機能がわかっても表現型との結びつきをうまく示すことが出来ないと言うことが起こりうる。そういう事態を防ぐために、前もって野生型植物や培養細胞を用いて様々な調査を行なっておくのが望ましいと思われる。私はこの変異体の仕事を始める前に、「セルロース合成阻害剤に耐えられるようになった培養細胞」を作成し、それを用いた研究も行っていた。それが役に立っている。また、マップベースクローニングを本格的に始める前に変異体の様々な生理学的性質を調べて、変異が引き起こした表現型についてある程度知ることが出来ていた。
「シロイヌナズナの変異体と、その原因遺伝子の単離」を行う利点として、「比較的体力を使わずにすむ」ということもあげられる。シロイヌナズナのマップベースクローニングは、作業は面倒だが普通の部屋ですべてが済む。またDNAは安定に保存でき、いつでも好きなときに分析できる(酵素のように失活することがない)ので時間に余裕がない時期でも少しずつ進めることが出来て便利である。結果もメンデルの法則に基づいて理屈通りに出てくるので安心して出来る。しかし、そのうちにまた「どうしても、ハードな生化学的な手法を用いなければこの問題を解くことは出来ない(翻訳後修飾とか)」ということも増えてくるかもしれない。
このデータは、広島大学自然科学研究支援センターの「DNA塩基配列決定サービス」によって得られたものである。この研究支援システムがなかったら成功することは無理だっただろう。各大学、研究所での研究支援体制をもっと充実、確立していただければ大変有り難い。
「セルロース合成阻害剤 (DCB) に耐えられるようになった培養細胞」を用いた自分の研究成果から、DCB を用いることで、非常に興味深いことがわかるに違いないと確信を持てるようになった。
それらの成果(二つの論文)は、内容も少なく引用されることもあまりない。しかし自分にとっては、確信を持てるようになったことで非常に意味があった。他の人から見るとそう思えないだろうが、一応このような論文を出すことも出来たので、間違いではなかったと思っている。

上の写真の右に示したのが、「セルロース合成阻害剤 (DCB) に耐えられるようになった培養細胞」 である。固まりを作りやすいのが特徴である。わたしはずっと「ペクチンが増えているので細胞同士がくっつきやすくなっているのだろう」と単純に考えていた。しかし、固まりを作ると言うことには別の意味もある。固まりの内部の細胞には酸素が届きにくい。そのことによって、内部の細胞は嫌気的な代謝をしているはずである(ミトコンドリアが通常の好気代謝をしにくい)。このことは、私がシロイヌナズナで見つけた変異体を調べてわかったことと共通している。代謝が嫌気的に変化すると、セルロース合成阻害剤 (DCB) に耐えられるようになるらしい。そのことは培養細胞とシロイヌナズナ個体で共通しているらしい。
シロイヌナズナの根の場合でも、表皮細胞やその直下の細胞はDCB処理でよく肥大するが、維管束に近い細胞、維管束の細胞は全く肥大しない。植物でも、組織の内部の細胞は酸素の供給量は低い。血管があるわけではないので自然拡散だけで酸素は供給される。低酸素状態に近い状態にある(と一時は思っていたが、よく考えるとシロイヌナズナの根はとても細いのでそんなことはないような気がする。また維管束は師管、導管液から栄養分と酸素が供給される)。
様々な変化(固まりの形成、ミトコンドリアの機能低下) → 低酸素条件とある程度似たなんらかの変化→ ? →DCB処理による影響(生育抑制、細胞肥大)が起きにくくなる: 同時に細胞壁等に様々な顕著な変化が起きる
というように最初は思っていたが、しらべていくとまた違う仮説が出てきたりもする。様々なことが複雑に相互作用しているので、正しい方向に研究を進めることが難しい。そこを何とかしなくてはいけない。
私は現在の職に就く以前、植物ホルモンの研究をしていた。その頃の考え方は
ホルモン → 受容体 → 因子A → ・・・ → 遺伝子発現
というような、直線的なものであった。
しかし今では、そのような直線的な仕組みは、あるとしても例外的なものであるようである。
様々な因子がつながりあったネットワークがあり、それに対するインプットがホルモン、アウトプットが遺伝子発現と考えた方がよいように思われる。インプット、アウトプットが複数あるのが当たり前のように思える。
というように考えていたが、植物ホルモンのシグナル伝達は予想よりも簡素、シンプルであることが明らかにされてきている。
シンプルな植物ホルモン http://www.rikenresearch.riken.jp/jpn/research/6122 理研、植物科学研究センターのすばらしい業績
シンプルな植物ホルモンが複雑多様な作用をもたらすということには、ホルモン同士の相互作用が重要であると考えられている。 それだけでなく、ホルモンで制御される多様な標的(例えばミトコンドリア、細胞表層など)のネットワークにも、複雑さをもたらす仕組み、因子があるということも考えられる。考えるだけではしょうがないので、何か糸口となることを見つけないといけない。
複雑なネットワークを数学的に研究する理論に、グラフ理論がある。秋山仁博士が専門にしていることで有名である。様々な応用ができるのが特徴であるらしい。このような分野の成果を、様々な応用例を参考にして取り入れられればよいかもしれない。
リンク
「最小コストフロー問題」というものが紹介されている。細胞内には様々なネットワークがある。代謝ネットワークでは代謝産物の流れがある。シグナル伝達ではホルモンや環境条件の信号がネットワークを流れる。生物は常に最小の資源で最大の成果(繁殖、増殖)を成し遂げようとしている。細胞内のネットワークでは、流れに必要なコストを最小にするように物質や情報が流れていると考えることもできる。
タンパク質同士が形成する反応ネットワークの構造が、刺激に対する頑健性 robustness に関わっているという話があった。
Structural Sources of Robustness in Biochemical Reaction Networks Science 12 March 2010: Vol. 327. no. 5971, pp. 1389 - 1391 Guy Shinar1 and Martin Feinberg2,*
この論文では、ある細胞内反応(EnvZ-OmpR system)のネットワークをグラフ理論で分析している。それを robustness と結びつけている。
「状態空間表現」というモデルが制御理論の分野で発達している。「複数の入力と出力を持つシステムをコンパクトにモデル化できる」そうである。システムの状態を、いくつかの、お互いに無相関な状態変数を組み合わせて表現する。PLS 回帰と似ている?
PLS 回帰では、「潜在的な因子」を抽出して予測精度のよいモデルを作る。「潜在的な因子」に対して何も知見がなくても、与えられたデータから自動的に抽出される。潜在的な因子を組み合わせることで出力を予測できるように計算する。「説明される変数と説明変数の間の共分散の最大化」を指標にして軸を決定する。
主成分分析を行うとお互いに無相関な主成分を求められる。それらを用いて重回帰を行う主成分回帰という方法がある。 単に線形に主成分を組み合わせるだけでなく、もっと複雑に組み合わせるようにすれば「状態空間表現」 のようになるかもしれない。
生物学では mRNA量を測定したり、酵素活性を測定したりして細胞のシステム、状態を推定しようとする。特徴として、得られる値はお互いに相関が高いことが非常に多いと言うことがある。多変量解析の方法を用い、お互いに無相関な複数の値(軸、次元、潜在的な因子)に再構成した方が数値として扱うには適しているのかもしれない。その場合、得られた軸が生物学的にどんな意味を持っているのかわかりにくくなる。しかし、それぐらいの段階に持って行かないといつまでたっても複雑だ、理解できないというままになってしまうのかもしれない。PLS回帰のような方法で得られた「潜在的な因子」の生物学的な意味をうまく導き出すというようなことが必要なのかもしれない。
統計を専門とされる先生が主成分分析の有用性を語っている。 http://www.spss.co.jp/success/opinions/files/031.html 成蹊大学 理工学部 岩崎教授 「主成分軸の(自分勝手な)解釈も楽しいことこの上ない。」と書かれている。主成分分析などで得られた結果をうまく解釈するところに、研究者としての腕の見せ所があるのかもしれない。私も多変量解析をよく勉強して、生物学的なデータから有用な「構造」「規則性」を見つけたいと考えている。
Cytoscape という、ネットワークの描画、解析のための素晴らしいソフトウェアが開発されている。 http://cytoscape.seesaa.net/ Excelによるネットワークとアトリビュートデータファイルの作成ができる。私のような素人がデータをそろえる際は Excel を用いる。Excel で作成したデータがそのまま使えると素人にとっては楽になる。http://cydoc.sourceforge.jp/cydocwiki/ に、素晴らしい日本語による解説文書が集積されている。
社会学、経済学等でも複数の因子の間の因果関係を明らかにすることは重要である。「因果序列図」 「グレンジャーの因果性テスト」 等のキーワードで、複雑な因果関係のネットワークを解明した例が紹介されている。多次元の時系列データを多数必要とするらしい。「時間的に先行するかどうか」ということは、因果関係を推論するうえで重要になる。
問題点としては、「雷光と雷鳴」のような場合がある。遠くに雷が落ちるとき、まず 雷光が光り、しばらくしてから雷鳴が聞こえる。「時間的に先行する」ことを指標に すれば「雷光が原因で雷鳴が起きる」と考えてしまうかもしれない。しかし本当の原因は放電現象であり、雷光も雷鳴も放電現象の結果同時に起きるものである。時間差は光速と音速の差によるもので、雷光が原因で雷鳴が起きる訳ではない。
生物学だったら、マイクロアレイ解析等を時系列的にかなりの回数行う必要があるだ
ろう。時間間隔、必要なデータ個数はやってみないとわからないだろう。
リンク1
時系列でなくても、「ホルモンの濃度を 0, 0.1, 0.2, .... と細かく上げていき、それぞれの条件で各因子の発現量を測定したデータを採取する」ことで、ホルモンに対する各因子の因果関係のようなものがわかるかもしれない (低濃度のホルモンに応答するものは、高濃度でのみ発現するものよりもホルモンの作用を一次的、より直接に受けていると仮定すれば)。
Baskin and Bivens らの論文 (1995) が、スクリーニング、セルロース合成阻害剤感受性の検査、成長測定を行うのに大変参考になった。彼らは単純な実験しか行っていないが、非常によい論文だと思う。
Baskin TI 博士は、植物の細胞表層、細胞形態決定に関わるすばらしい業績を次々と上げている偉い人である。
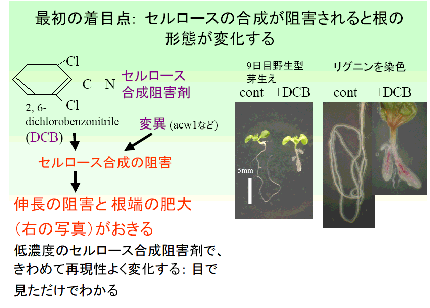
Baskin and Bivens の論文にあるように、セルロース合成が阻害されると根が肥大する。微小管重合を阻害する阻害剤(オリザリン)でも根が肥大する。Baskin 博士が書いているようにそれらの二種の処理による肥大は、見かけが非常によく似ている。そのことを、下の図のように解釈した。
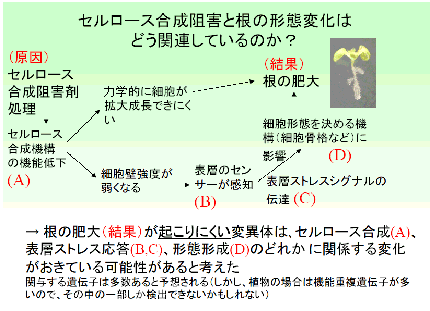
「根の肥大が起こりやすい変異体」は、すでにいくつも見いだされていたが、「おこりにくい」ものを見つけようとした人はいなかった。
「おこりにくい」変異体は、図の (A)(B)(C)(D) のどこかに関係している遺伝子が変化しているのではないかと考えた。このように関与している遺伝子が多数ありそうなのはあまり望ましいことではないかもしれない。しかし、植物では機能が重複している遺伝子が多いので、その中の一部しか検出されないかもしれない。また変異したことで生育がきわめて悪くなるものは、扱うのが難しい。あまり候補を絞るような条件では、何も出てこなくなるかもしれないし、出てきてもうまく育たないものばかりになるかもしれない。私の場合自分が考えて自分でやるのだから、どんな結果になっても文句を言われることはない(他の研究者と比べて、ずっと高いリスクをとれる)。とにかくやってみようと考えた (もちろん上に書いたように、野生型や培養細胞を用いて様々なデータを得ていたので「うまくいくだろう」という確信はあった)。
実際には、「根の肥大がおこりにくい」以外にも興味深い変異形質を同時に示すものが見つかった。運が良かっただけかもしれないし、このスクリーニングが何らかの非常に重要な分子機構の変動を敏感に反映しているのかもしれない。また、セルロース合成という非常に基礎的で大量の糖を消費する代謝は、他の基礎代謝と密接不可分な関係にあるということを示しているのかもしれない。
2009年9月に、シロイヌナズナの代謝経路全体をモデル化し、その flux を線形計画法(モデルは平衡状態で、Flux の総和を最小にする)によって分析した論文が発表された。細胞壁に流れる代謝産物が主要な因子になっている。細胞壁(セルロース合成)が、基礎代謝における糖の flux を考える上で必須の成分であることを示している(その割には無視されていることも多かったような気もする)。
A Genome-scale Metabolic Model of Arabidopsis thaliana and Some of its Properties. Poolman MG, Miguet L, Sweetlove LJ, Fell DA. Plant Physiol. 2009 Sep 15. [Epub ahead of print]
また、このスクリーニングは「セルロース合成が阻害されたことによる形態変化を、打ち消す変異のスクリーニング」と言うこともできる。「サプレッサー変異」のように解釈することもできる。ふつうは「suppressすべき変化」は何らかの遺伝的な変異によって引き起こされる。しかし、このスクリーニングのように阻害剤を用いることもできる。セルロース合成阻害剤は非常に低濃度でよく効き、培地中で安定なのでこのようなことができる。
「サプレッサー変異」を取ろうとしても、suppress するべき変異が強すぎてサプレッサーが全然取れないということもあり得る。阻害剤を使った場合は、阻害剤の濃度を適当な濃度(効果がある濃度で、できる限り低くする)にすることで作用を調節できる。それによって「サプレッサー変異」に相当するものが取れやすくなるという可能性もある。このことはかなり重要なことらしい。
しかし阻害剤がよい性質を持つ(特異性が高く低濃度でよく効き培地に加えたままでも分解されにくい)ものでないと都合が悪い。
除草剤は野外環境で安定でないと実用にならない。植物病原菌が作る毒素も、不安定ですぐに失活するものなら植物にダメージを与えることはできない。既に数多く見いだされている除草剤や病原菌由来化合物は、変異体のスクリーニングに使うためによい性質を備えている可能性が高い。
「サプレッサー」だけでなく「作用を増強する変異」も取ることができる。セルロース合成阻害剤についてもそのようなものを見つけることができる。それらの多くはセルロース合成に関わる因子(CesA など)の変異だと予想される。
DCB は約100nM, IXB は 10nM 以下でも根の形態を大きく変化させることができる。 しかしそれらの濃度は培地の組成、育成条件によって大きく変わる。できる限り低濃度で効くような条件を見つけておけば、薬剤をほんの少し入れれば済むようになる。そのほうが特異性が高く、経済的で安全性も高まる。
植物ホルモンの場合も、ウニコナゾール等の生合成阻害剤が、変異体の単離に有効に用いられている。南原博士らによって、ウニコナゾールを用いたアブシジン酸関連変異体のスクリーニングに関して丁寧に説明されている。 植物ホルモン応答性が変化した突然変異体のスクリーニング法 植物化学調節実験マニュアル(植物化学調節学会、吉田茂男・編 1997年発行)112ページから
今回のスクリーニングでは、培地組成が「1 x MS salt, 3% ショ糖」が主成分である。これは BY2 の培地をそのまま使ったものである。「セルロース合成阻害剤 (DCB) に耐えられるようになった培養細胞」を調べていた経験から決めた。 また Baskin らの論文でも、ショ糖を 3% 加えている。理由ははっきりとはわからないが、ショ糖が多いと根は良く肥大する。またセルロースの蓄積量も減りやすくなる。このことは以前から他のグループのセルロース関連変異体の論文にも書かれている。 培地組成はいくらでも変えられるので、どんな組成がよいかは事前の様々な研究例、自分の実験結果から適当に判断するしかない。そこでそれぞれの研究者の経験や知識による違いが試されるのかもしれない。異なる培地組成でスクリーニングすれば、また異なったタイプの変異体が見つかるかもしれない。
低酸素条件、ミトコンドリア関連変異などの要因により基礎的な代謝が切り替わることは、細胞にとって非常に重大で大幅な、様々な変化をもたらすらしい。
動物の場合、腫瘍が固まりを作る場合がある。固まりの内部の細胞の代謝は嫌気的になる。そのことによって、ミトコンドリアのアポトーシスに関わるタンパク質の活性に変化が生じ、抗腫瘍薬が効きにくくなると言うことがあるそうである。動物の腫瘍では解糖系代謝が亢進することが知られている(Warburg, ワーバーグ(ワールブルグ)効果)。これは「中心部が低酸素に陥っている」だけではなく、別の意味があるということが示されている。
http://www.geriatric.kuhp.kyoto-u.ac.jp/top/s_cel.html 京都大学医学部の近藤先生が書かれた解説: 近藤先生は、解糖系代謝が亢進すると酸化ストレスが軽減されることを示している。このことが他の生物でも成り立つのかどうかは、わからない。
Science 2009年5月22日号 Volume324 Number5930 に、「ワールブルグ効果とは何か:細胞増殖の代謝要求
注)ワールブルグ効果:がん細胞において、酸素が十分供給されていても嫌気性の解糖系代謝が亢進している現象」
Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation
Matthew G. Vander Heiden, Lewis C. Cantley, and Craig B. Thompson
という Review が掲載された。
http://www.cosmobio.co.jp/aaas_signal/archive/pp_20091117_1.asp PKM2のチロシンリン酸化とグルタミン代謝がワールブルク効果の別の見方を提示する
ペントースリン酸経路 Pentose Phosphate Pathway は、Glucose-6-phosphate から始まって Fructose-6-phostase と Glyceraldehyde-3-phosphate を生産する。その過程で NADPH を生成する。NADPH は Glutathione reductase の補酵素となって還元型のグルタチオンを生成する。植物にもグルタチオンペルオキシダーゼがあって、過酸化水素を消去するのに働く。
解糖系とペントースリン酸経路は一体となってNADPH の生産に働く。ミトコンドリアの代謝ではATPが効率よく生産されるがNADPH の生産は起きにくい。細胞増殖にはATPとNADH, NADPHのバランスが重要であるらしい。ATP だけが作られても、増殖に必要な生体分子を合成することができにくい。解糖系とペントースリン酸経路を活性化することでNADPH の生産を高めることが、増殖速度を高めるのに働いていると言うことを書いた解説があった。
植物、真菌、放線菌などでは二次代謝系が働いて色素などを大量に合成することがある。二次代謝経路では NADPH を消費する反応が多い。二次代謝産物を多く作る細胞は、細胞分裂を行っていないことが多い。よく増殖する培養細胞は二次代謝産物を作りにくいことが多いことも知られている。NADPH の取り合いのようなことがあるのかもしれない。
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=pubmed&cmd=Retrieve&dopt=AbstractPlus&list_uids=16728594&query_hl=1&itool=pubmed_DocSum 動物のワーバーグ効果には p53 という非常に重要で多機能な転写因子が関与し、ミトコンドリアの呼吸系を制御している。
「解糖系」や「ミトコンドリア」は、きわめて基本的な研究課題で長年の蓄積があるが、まだよく知られていない現象や知見がたくさんあるのかもしれない。そしてそれらの未知の現象は細胞、生物にとって、とても重要な因子であるのかもしれない。 今後「解糖系」や「電子伝達系」の研究が見直される、再注目されるかもしれない。
動物での例が、サイエンス誌からのメールマガジンで紹介されていた。
Evidence for an Alternative Glycolytic Pathway in Rapidly Proliferating Cells
急激に増殖する細胞において解糖系の別経路の存在が明らかになった がん細胞をはじめ増殖する細胞は、栄養素を 効率よく取り込んでバイオマスにするために代謝を変化させる必要がある。今回、ホスホエノールピルビン酸(細胞内の ピルビン酸キナーゼの基質)が哺乳類の細胞内でリン酸供与体として働きうることが示された。 Matthew G. Vander Heiden, Jason W. Locasale, Kenneth D. Swanson, Hadar Sharfi, Greg J. Heffron, Daniel Amador-Noguez, Heather R. Christofk, Gerhard Wagner, Joshua D. Rabinowitz, John M. Asara, and Lewis C. Cantleyhttp://www.sciencemag.org/cgi/content/abstract/329/5998/1492
「バイオマスの効率的な生産」は、植物の遺伝的改良において非常に重要である。解糖系の研究、効率化が植物でも必要かもしれない。がん細胞の研究が植物の解糖系の効率上昇に役立ったりすれば、さらにそれによって植物由来のバイオ燃料の生産効率を高めることができたりすればとても興味深い。
ミトコンドリアの電子伝達系は、パーキンソン病と深い関わりがある。http://www.sciencemag.jp/highlight/index.jsp?pno=232 で紹介されている。 Translational Medicine 10月6日号: パーキンソン病患者の脳細胞ではミトコンドリアが機能していない Parkinson’s Brain Cells Abandon Mitochondria 。他の研究において、パーキンソン病患者では電子伝達系を構成する5つの複合体のうち1つの機能が失われていることが示唆されているが、Scherzer らは、1つだけでなく、実質的に、電子伝達系を成立させるためにミトコンドリアに不可欠なすべての構成要素が欠如していることを見出した。
Science 誌に、代謝研究特集が掲載された。 http://www.sciencemag.org/content/330/6009/1337.full.pdf INTRODUCTION Metabolism Is Not Boring Perspective、さらに review が4つ掲載されている。がん、Autophagy, 概日リズム、代謝工学について論じられている。
基礎的な糖代謝経路は、「メタボローム」研究や「代謝シミュレーション」研究の格好の対象である。
植物は正常な生育に必要な必須因子として働く多種類のホルモン類を合成している。それらの量が、基礎代謝の速度とリンクして制御されていることがあるかもしれない。
植物に有用物質 (3F: Fiber = cellulose, Food = starch, Fuel = Lipid) をたくさん作らせようと思ったら、基礎代謝経路について調べないわけにはいかないのではないか。
Fig. 1 は、Baskin and Bivens (1995) の論文を真似て、根の伸長と肥大に対する阻害剤やホルモンの影響を見たものである。変異体に対する様々な生理活性物質の影響を、比較的簡単に短時間で、定量的に調べて野生型と比較することが出来る。変異体について調べるのに適している。私はこの実験をたくさん行っていて、もっと多くのデータがある(細胞骨格の重合阻害剤の影響など)。しかし論文では余計なデータは省かなければいけない(読む人を混乱させる元になる)ので、必要な分しか出していない。
Fig. 3 にあるように、この変異体の初期成長 (post-germination growth) は、培地の糖濃度に強い影響を受ける。これは「代謝が異常なので、細胞内の糖濃度の調節がうまくいかずに、このような結果になる」という可能性がまず考えられる。しかし、「糖シグナルやホルモンに対する感受性が増大している」という可能性もある。
シロイヌナズナのゲノム配列が決定され、すばらしい使いやすいデータベースを自由に使えるようになった。そのため私のようなものでもマップベースクローニングができるようになった。マップベースクローニングは、非常に手間と時間がかかるが、それを我慢できる立場にあるならば基本的に難しいことはない。もちろん複雑、unusual な遺伝をする変異などでは難しいこともあるだろう。運悪く、目的の遺伝子座の周りで組み換えが非常に起こりにくいと言うこともあり得る。変異体が示す形質が弱かったり、安定しないと研究が大変難しくなる。私の場合は、単純な劣性変異で、変異体と正常なものの見分けも簡単、確実に安定して行えた。遺伝子座の場所を詰めていくのも、ものすごく困難というほどではなかった。そのため何とか成功することができた。
植物細胞工学別冊「モデル植物の研究プロトコール」(秀潤社) を熟読して参考にさせていただいた。
・ 変異体 (Col-0 に由来する) と野生型 (Ler) を交配し、種子を得た。さらに育成し、1:2:1 に分離する種子をたくさん収穫した。
・ それらを、セルロース合成阻害剤を含む培地に播いた。変異形質を示す芽生えを選択した。私の場合はシャーレ内の寒天培地で済むので楽だった。
・ 選択された芽生えから、それぞれ DNA を抽出し保存した。 抽出法は、以前は「モデル植物の研究プロトコール」に書いてある簡易法を用いていた。このDNAの取り方では、プライマーによっては PCR がうまくいかないことがある。現在はもう少し手間をかけた方法にしている(酢酸カリウムでSDSと変性タンパク質を沈殿させた後上清をイソプロ沈殿してTEに溶かす)。
抽出バッファーには、DNA の沈殿を促進するためにリニアポリアクリルアミドを加えると都合がよい。 Ethanol precipitation of DNA with linear polyacrylamide as carrier C Gaillard and F Strauss Nucleic Acids Res. 1990 18: 378

・ CAPS マーカーのためのプライマーを用意した。PCRを行い、制限酵素で切断し電気泳動でそれぞれのマーカーの場所における遺伝形質を調べた。
・ だいたいの位置がわかったら、そこに近いマーカーをさらに作成し、場所を限定していった。
モンサント社が公開しているSNPのデータをTAIR からダウンロードして、それに基づいて適当にプライマーを設計した。モンサントのデータはエクセルで読み込めるようになっている。SNP によって区別できる制限酵素のリストもすでに検索され書き込まれている。データの信頼性は非常に高い。
エクセルのオートフィルタで、作りたい場所のBACの名前を含む行だけを表示する。見つかったら、カラム M, N に書いてある制限酵素のリストを見る。都合の良い酵素が使えるSNPが見つかったら、カラム K に書いてある flanking sequence をコピーする。
TAIR のホームページの、 seqviewer のテキストボックスにペーストする。その配列を含む部位を検索して nucleotide sequence view を表示する。 flanking sequence は赤地に白の文字で表示されている。SNP の部分を中心として上下300bp ずつコピーする。
それを WebPrimer http://seq.yeastgenome.org/cgi-bin/web-primer にペーストしてプライマーを設計する。うまくいかないときは場所を少し変えてみる。
設計できたら、増幅される配列全体を Webcutter http://rna.lundberg.gu.se/cutter2/ にペーストする。CAPS に使う酵素で切れる場所を確認する。SNP によって切断箇所が変わっても、断片の大きさの変化が小さい場合がある(酵素が4塩基認識の場合に多い)。その場合は使えないので、そのSNPはやめて、別のSNPで設計をやり直す。
目的の遺伝子座に近くなってくると都合の良いSNPが存在しなくなる。その際は制限酵素で切って泳動するのではなく、PCR産物をそのままシーケンスすることでSNPを判定した。
・ 最終的には、限定された領域内に存在した遺伝子 (約30個)について、読み枠の塩基配列を決定して変異が見つかるまでそれを続けた。「こんな遺伝子が関係あるはずないだろう」とずっと思っていたものが、本物だった。要するに全然当たりが出なかった。ある日「この変異体の性質は、ミトコンドリアに異常があると考えればすべて説明できる」ということに気がついた。それに長い時間がかかった。ミトコンドリアと関連する遺伝子は領域内にこれしか残っていなかった。変に予想せずに領域の中央にある遺伝子から順番にひたすらシーケンスした方がよいかもしれない。そうでないと自分の考えが間違っていたときに時間をかなり無駄にしてしまう。
Fig. 7 に書いたように、この変異体では細胞全体(細胞壁、細胞質、オルガネラ)に渡って顕著な代謝変動が起きている。それによって、ミトコンドリアのNADH代謝の欠陥を補償(complement)している。 私はこの現象を「植物の代謝可塑性」と呼んでいる。論文を書いたときにはそこまで考えを進めることは出来なかったので、論文には書いていない。しかし代謝が可塑的なのは植物に限ったことではないので、あまりそのことを強調しても仕方がないような気もする。
植物の細胞壁構成成分がかなり変化しても細胞は生育できることが多い。代謝的な可塑性が高いと解釈することもできる。
大腸菌がアミノ酸飢餓状態に陥った際、緊縮応答と呼ばれる応答が起きる。その際様々な代謝変化が起き、同時に細胞壁合成が抑制されることが明らかにされている。細胞内の栄養源量を増やすために、細胞壁合成に割り当てる資源を節約するということは普遍的な仕組みなのかもしれない。
最近「合成生物学 Synthetic biology」「合理的ラショナルゲノム設計 Rational Genome Design」 という概念が注目されている。細胞内に新しい代謝経路を構築するように、遺伝子を設計しバクテリアや植物に導入する。それによって有用代謝産物の増産などの機能改良を人為的、設計通りに引き起こす。電子機器を作るために電子回路を設計するように、新規の代謝回路を設計する。設計した結果細胞内代謝がどう変化するかを、コンピュータ内にモデルを組みシミュレートすることができる。 参考になる資料 http://crds.jst.go.jp/output/pdf/09sp11.pdf 「環境適応型作物のゲノム設計技術」
「細胞で起きることなんて、あまり予想通りになっていることなんかないんだから余計なことを考えずに変異ライブラリーを作ってそこからスクリーニングした方が予想外、想定外のものが見つかって面白いんじゃないか」という気もしなくもない。しかしいつまでもそういうことをしているわけにもいかないのだろう。網羅的測定技術、ゲノム配列と発現のデータベース、シミュレーション技術の進歩(情報生物学の成果)が「代謝回路設計」の基盤として用いられている。そこに新しさがある。
「将来のゲノム設計士を育成する国際科学技術コンテストをウェブ上で初開催」というイベントが2010年5月から開かれた。 http://www.riken.jp/r-world/info/info/2010/100524/index.html
代謝の改良を考える際に、NADHの代謝はきわめて重要だろう。電子伝達と関係があると言うことで「細胞内の電子回路」といえるかもしれない。私が見つけた変異体は、NADHの代謝に関する変異ということで、NADHを反応に使用する代謝を人為的に強化するように設計するための親株として使えるかもしれない。
酵母 K. lactis 由来のアルコール脱水素酵素(KLB)はNADHを補酵素とする。この酵素を水素細菌に導入し、ヒドロキシアセトン Hydroxyacetone をプロパンジオールという誘導物質に変換する機能を強化することが成功している。 (1) http://www.nedo.go.jp/iinkai/kenkyuu/bunkakai/18h/jigo/45/1/5-5.pdf
(2) http://www.nedo.go.jp/iinkai/kenkyuu/houkoku/20h/chukan/17-4.pdf
(1)の資料の9ページに、説明がある。水素細菌のヒドロゲナーゼによって NAD から NADH の再生を行い、NADH を KLB の補酵素として用いている。私が見つけた変異体は、NADHの代謝に関する変異ということで、ADH, NR などの酵素が働きやすくなっているかもしれない。実際に起きている代謝変化も、それで説明できるように FIG7 には書いた (NR、アラニンの部分)。しかし実証はされていない。
vim: set ts=8 sts=2 sw=2 et ft=a111_modified_flexwiki textwidth=0 lsp=12:
![]()