現在の研究テーマ
生物種によらない膜交通機構の基本原理の追求
膜蛋白質やオルガネラ内腔タンパク質、さらに分泌タンパク質 (以降、これらを積荷タンパク質と表現します) は、小胞体膜上で合成された後、シスゴルジ囊に送られ、ゴルジ槽成熟の結果、トランスゴルジ網 (TGN)に到達します。TGNは積荷タンパク質の選別の中枢であり、様々な細胞膜ドメインやオルガネラへ輸送されるポストゴルジ小胞が形成され、その内部へとタンパク質が選別されて詰め込まれる、と考えられています。しかし、このTGNにおける積荷タンパク質の選別とその後のポストゴルジ輸送の分子機構については、まだ詳細は不明です。ポストゴルジ輸送はオルガネラの機能維持や細胞膜ドメインの形成・維持に必須な機構であり、生命機能の維持には不可欠な過程です。実際その破綻は、微絨毛封入体病・多発性嚢胞腎・網膜変性症などを含む様々な疾患を引き起こすことが報告されています。つまり、ポストゴルジ輸送の分子機構解明は、細胞生物学の基本的命題であるとともに、その解明により極性輸送欠損が引き起こす様々な疾患についての治療法の開発に直接つながることが期待されます。
ショウジョウバエ視細胞を用いた極性輸送研究
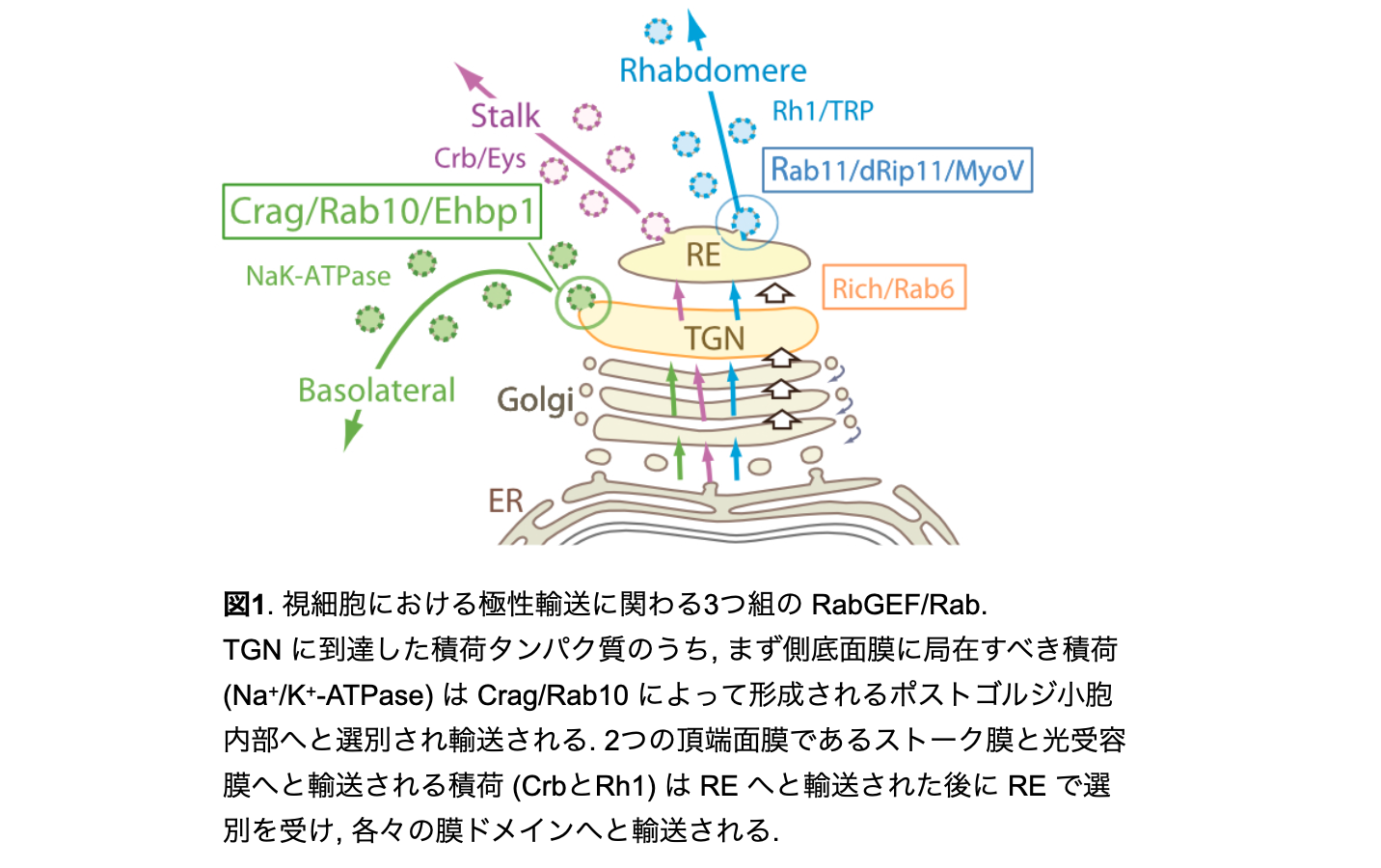
佐藤研究室では長年、明瞭な極性をもつショウジョウバエ視細胞をモデルとした極性輸送の研究を行ってきました。この過程で、ゴルジ体のトランス側では3つのRabGEF/Rab が相互にずれ合いながら局在し、特定の膜ドメインへ極性輸送を制御することが明らかにしてきました (図1)。特に、Pcs/Rab11はハエ視細胞ではゴルジ体の最もトランス側と光受容膜基部のポストゴルジ輸送小胞に局在し、Rh1を光受容膜へと届ける役割を持つことを示しましたが、Pcs/Rab11は本来エンドサイトーシス経路で機能する因子であり、リサイクリングエンドソーム(RE)マーカーとして用いられています。そこで、ゴルジ体とREとの関係についての研究を開始しました。
TGNとREの接着と解離と選別輸送
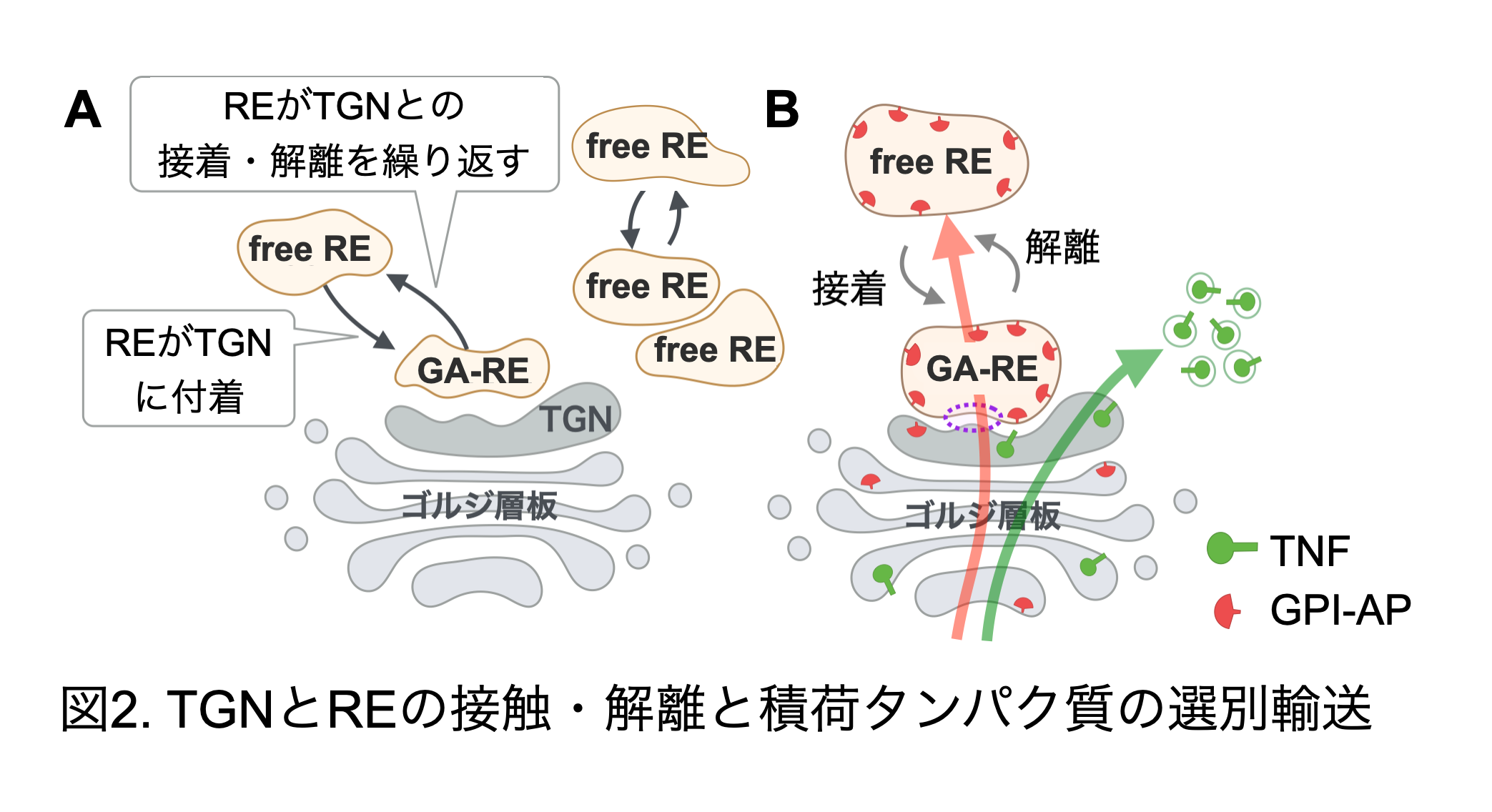
ゴルジ体は生合成されたタンパク質の成熟と選別を行うオルガネラで、シス槽・メディアル槽・トランス槽・TGNが積み重なった層板構造を基本単位としています。一方、REは細胞膜からエンドサイトーシスによって細胞内に取り込まれた物質を再び細胞膜へと戻す役割を持っています。両者は独立した異なるオルガネラと考えられています。私達は、ハエの様々な組織、微小管破壊によりゴルジ層板を散在させた哺乳類細胞やウニにおいて、ゴルジ層板とREの関係を検討した結果、REはゴルジ層板付着型 (Golgi attached-RE: GA-RE)と遊離型 (free-RE)の2つの状態で存在することを見出しました (図2A)。さらに哺乳類やショウジョウバエ培養細胞をライブイメージング観察すると、この2つの状態はREのゴルジ層板に対する付着と遊離の繰り返しによって生じていた(図2A)。植物細胞ではREは定義づけられていないが、ゴルジ体の一部と考えられているTGNがゴルジ体のトランス側への接着と解離を繰り返していることが報告されている。研究代表者らの見出した動物細胞におけるREの挙動は、この植物細胞のTGNの挙動と酷似しており、植物TGNと動物REが極めて近い関係にある細胞小器官であることを示している(図ではTGNとREを分離して示した)。次に、HeLa細胞を用いて積荷タンパク質輸送とGA-REの関係を検討した。その結果、積荷タンパク質の種類によってゴルジ層板/TGNからGA-REを経由して細胞膜へと輸送されるものとGA-REを迂回するものとが存在することがわかった (図2B)(Fujii et al., 2020 J Cell Sci. Feb; Fujii et al., 2020 Commun.Integr.Biol.;及び未発表データ)。この結果はゴルジ層板/TGNとREの境界で積荷タンパク質の選別が行われ、一部の積荷タンパク質をREに送り出していることを示している。つまり、ゴルジ層板/TGNとREの境界が、異種の膜系間の境界領域における積荷選別の重要なモデルとなると考えられる。