
真核生物のゲノムは、染色体テリトリーやTopologically-associated domain (TAD) などの階層的なクロマチン構造によって組織化されています。その物理的制約は、同じTAD内のエンハンサーやプロモーターなどの調節配列の接触頻度を高くし、TAD外の領域とは接触頻度を低下させることで、適切な遺伝子発現制御に寄与していると考えられています。脊椎動物のCTCFはこのクロマチン構造を制御するための重要な因子であり、遺伝子の転写制御や染色体の三次元構造形成に関与しています。実際CTCFがTADの基部に結合し、コヒーシンと共同して染色体ループ構造の形成に関与することが知られています。CTCFはインスレータータンパク質としてもよく知られているため、ウニのインスレーターの研究をする我々も注目していました。そこで、ウニの初期発生におけるCTCFの機能を解析するために、ウニのCTCFホモログ(HpCTCF)をクローニングし、その構造および機能の解析を行いました。
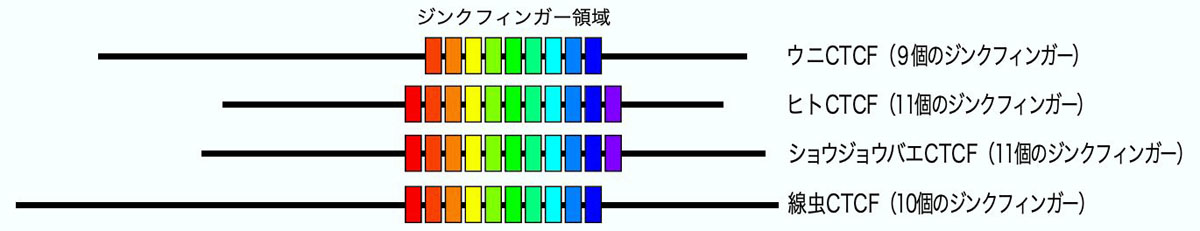
その結果、哺乳類のCTCFは11個のジンクフィンガーをもつことで知られていますが、
ウニのCTCFには9個のジンクフィンガーしかありませんでした。アミノ酸配列を詳細に解析した結果、これらのジンクフィンガーは、哺乳類の2番目から10番目のジンクフィンガーに相当するものであることがわかりました。さらに複数のフィンガーが、旧口動物(ショウジョウバエや線虫)のCTCFがもつジンクフィンガーとの高い相同性を有していました。よって
ウニのCTCFは、新口動物と旧口動物の共通祖先がもっていたであろう進化の歴史古来の基本的な機能を保持していると予想されました。しかもこれらのフィンガーは、哺乳類において細胞増殖に関連した機能が示唆されていました。
また、ウニの発生過程におけるHpCTCFの発現部位を調べたところ、HpCTCFは胚全体に発現しており、しかも局所的に細胞が増殖する植物極板の周辺で強い発現が観察されました。さらに、HpCTCFとGFPの融合タンパク質(HpCTCF-GFP)をウニ胚に発現させてその挙動をタイムラプス解析したところ、ウニのCTCFは間期の核内に均一に分布しており、分裂期に入ると消失し、分裂後に再びCTCFが核に集合することがわかりました(左動画)。
さらに、HpCTCF-GFPと一緒にヒストンH2BとmCherryの融合タンパク質(H2B-mCherry)をウニ胚に発現させて、細胞周期の進行にともなう挙動を詳細に解析したところ、HpCTCFは間期の核内に均一に分布しており、分裂期に入って染色体が凝縮するとき(分裂前期)に減少し、分裂中期と後期の染色体上には検出されませんでした。そして、分裂の終期に間期の核を再構築する過程でCTCFは核に集合し始めることがわかりました(右動画)。
さらに、HpCTCFの機能を解明するために、モルフォリノアンチセンスオリゴ(MO)によるHpCTCFのノックダウンを行ったところ、桑実胚期〜胞胚期で発生が停止しました。左のタイムラプス動画では、上列でHpCTCFをノックダウンするMO(HpCTCF-MO1)を顕微注入した胚で、下列はコントロールのMOを顕微注入した胚です。下列のコントロールMO注入胚は胞胚期まで正常に発生し胞胚腔内もクリアでしたが、HpCTCF-MO1注入胚は徐々に胞胚腔が濁っていき、無秩序に細胞が詰め込まれたような状態となりました。通常の実験条件ではこのような状態で発生が停止しましたが、タイムラプス解析では左の動画のようにその後胚が崩壊していきました。

HpCTCF-MO1注入胚で発生が停止したタイミングでDAPIまたはHoechstによる染色をおこなったところ、コントロール胚では多くの細胞が丸い間期核の状態でしたが、HpCTCF-MO1注入胚(CTCFノックダウン胚)では多くの細胞で染色体が凝縮したままの状態でした(右図)。しかも、この凝縮染色体のほとんどがヒストンH3のSer10がリン酸化されていなかったことから、分裂期の終期で停止していたことが明らかになりました。
さらに、HpCTCF-MO1と一緒にH2B-GFP mRNAを注入してタイムラプス解析による詳細な解析を行いました。その結果、HpCTCF-MO1注入胚も桑実胚期までは正常な分裂像が観察されましたが、桑実胚になってから分裂期の後期で姉妹染色分体が糸を引いたような状態で完全には分離できず、その後異常な形態の核が形成される様子が観察されました(左動画)。
しかし、ウニのCTCFをCRISPR-Cas9システムを用いたゲノム編集によりノックアウトしたところ、高い変異率は得られたものの、発生に異常はみられませんでした。少なくとも受精後10日のプルテウス幼生までは、正常に発生しました。つまり、ウニCTCFは初期発生における分裂期の進行には重要ですが、その後の発生過程における遺伝子発現への貢献は低いと考えられます。
以上のように、脊椎動物のCTCFは間期の核で重要な役割を果たすとされてきましたが、ウニのCTCFは初期発生において分裂期の進行に必須であり、とくに分裂終期から間期への移行に重要な役割を果たしていることが明らかになりました。このような細胞増殖におけるCTCFの役割が、新口動物と旧口動物の共通祖先がもっていたCTCFの基本的な役割であることが、本研究により示唆されました。
