ほとんどの動物は外見上左右相称な形をしていますが、一見左右相称に見える体であっても体内の臓器は左右非相称に配置されています。このような左右非相称性を確立する分子メカニズムは長年研究されてきました。マウスでは、7.5日胚の腹部にnodeとよばれるくぼみが形成され、その中心部に存在する繊毛が回転運動を行うことにより左向きの水流が生じます。その流れを側方に形成されている繊毛が感知することで左右非相称性が決定します。一方カエルでは、H+/K+-ATPase イオンポンプの局所的な開閉による膜電位の変化が左右非相称の決定に必要であると報告されています。脊索動物のホヤでは、神経胚期に形成される繊毛が胚自体が回転させ、その胚の回転が止まったときに卵膜と接している部位にNodalが発現し、この Nodalの非相称な発現により左右相称性が破れることが明らかにされています.
左右非相称性の決定
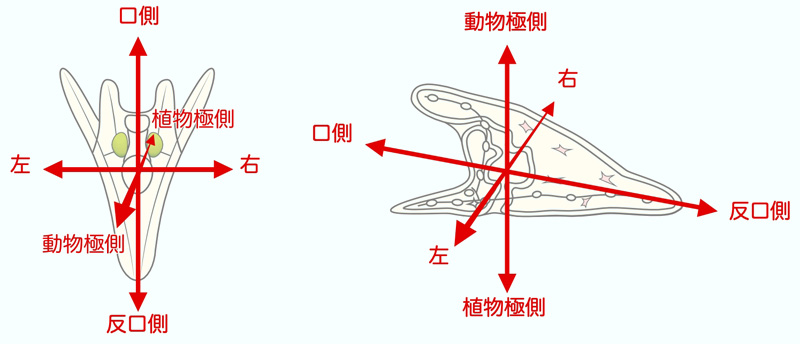
ウニの成体のからだは五放射相称ですが、プルテウス幼生は左右相称です。原腸陥入において、植物極からの陥入口(原口)が幼生の肛門となります。そして、原腸は動物極側に伸長しながら前方に傾斜し、外胚葉に達して形成される口陥が幼生の口となります。そこで口側(前方)を上にして、動物極側(背中側)からプルテウス幼生を見たときに、左にくるのが左側、右にくるのが右側と考えます。
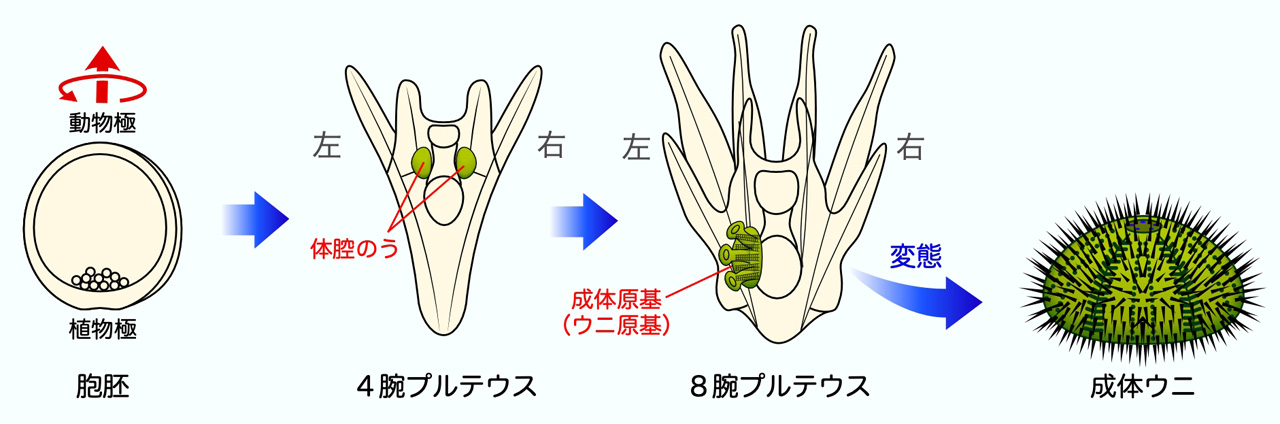
左右相称なプルテウス幼生には左右に一対の体腔のうが形成されますが、8腕プルテウス幼生になるとほぼすべての個体で右側の体腔のうが退縮し、左側の体腔のうのみが成体原基を形成します。一方で胞胚期のウニ胚は、ほぼすべての個体が繊毛運動により動植物軸を中心に植物極側からみて左回りに回転しながら遊泳します。この事実から、ウニ胚においても繊毛運動による胚の回転運動が左右非相称の決定と確立に関与しているのではないかと考えました。
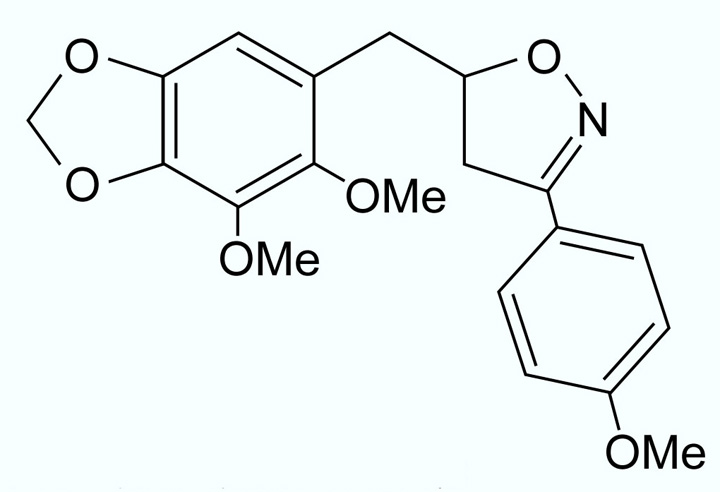
ウニ胚から繊毛を除去することが報告されていたDillapiol isoxazoline derivative 1 (DID1)という薬剤でウニ胚の回転運動を阻害したところ、右側または両側に成体原基が形成されるプルテウス幼生の割合が増加しました。また、同様の現象がウニ胚を寒天に封入して繊毛運動を阻害した場合にも観察されたことから、ウニ胚における左右非相称性の決定に繊毛運動が関与していると考えられました。そして更なる解析により、DID1が効果を示すのは受精後6〜8時間の桑実胚期を中心とした時間帯であることが判明しました。しかし、桑実胚期には胚の回転運動は起こらないため、胚全体の回転運動は左右非相称性とは関係ないことがわかります。
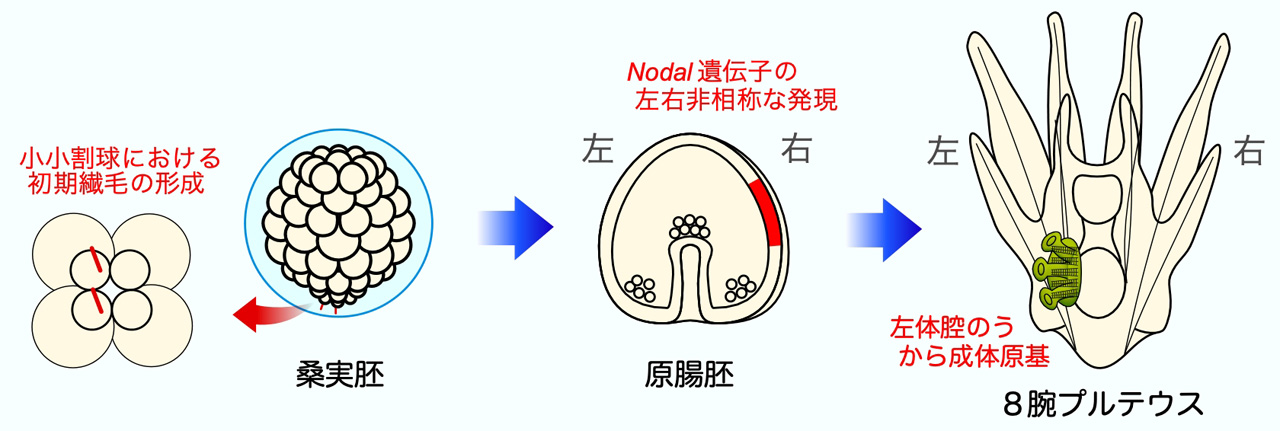
そこで次に、受精後6〜8時間の桑実胚で形成される繊毛を調べたところ、ちょうどこの時期に小小割球上に最初の繊毛が形成されることが明らかになりました。この初期繊毛が左右非相称性の決定に重要であると考え、現在さらに解析を進めています。また、初期繊毛からNodal遺伝子の左右非総称な発現までの調節経路も明らかにしていきたいと考えています。
